2023 © Erasmus MC, Rotterdam. Alle rechten voorbehouden. Niets uit deze uitgave mag worden verveelvoudigd, opgeslagen in een geautomatiseerd gegevensbestand of openbaar gemaakt in enige vorm of enige wijze, hetzij elektronisch, hetzij mechanisch, door fotokopieën, opnamen of enige andere manier, zonder schriftelijke toestemming van de uitgever.
Dit bestand is gepubliceerd op vrijdag 14 april 2023 15:24.
Manieren om het cerebellum onder te verdelen (zie vooral afb. 19.1 van Purves op p. 404 en Sobotta, afb. 12.72 op p. 253):
Vermis – hemisfeer (op basis van vorm)
Lobus anterior – lobus posterior – flocculonodulaire lob (op basis van lobulaire structuur van het cerebellum)
Vestibulocerebellum - spinocerebellum – cerebrocerebellum (op basis van verbindingen en functie)
Cerebellaire schors – cerebellaire kernen (op basis van oppervlakkige vs diepe structuur)
(eventueel) Zonale indeling (A-B-C-D-zones: op basis van verbindingen schors- kernen)
Laterale ventrikel (cornu posterius)
Cerebellaire kernen (nucleus dentatus)
Witte stof
Cerebellaire schors
Overgang verlengde merg/ruggenmerg
Cerebellaire pedunculi zijn grote vezelbundels die het cerebellum verbinden met de hersenstam. Onderscheiden worden:
Pedunculus cerebellaris inferior
Bevat aanvoerende vezels uit verschillende hersenstam gebieden en uit het ruggenmerg.
Pedunculus cerebellaris medius
Bevat aanvoerende vezels uit het de basale ponskernen (die op hun beurt weer informatie uit de grote hersenen ontvangen).
Pedunculus cerebellaris superior
Bevat afvoerende vezels uit de cerebellaire kernen.
Dus: pedunculus cerebellaris inferior en medius sturen informatie naar het cerebellum. Pedunculus cerebellaris superior stuurt informatie vanuit het cerebellum naar de hersenstam.
Noot: van de zgn. tractus spinocerebellaris ventralis wordt beweerd dat deze ook via de pedunculus cerebellaris superior het cerebellum bereikt (dus aanvoerende vezels). In principe kan deze tractus echter ook gezien worden als een deel van de pedunculus cerebellaris inferior. Dit maakt de indeling van de pedunculi in twee aanvoerende en één afvoerend simpeler.
Aanvoerende informatie bereikt het cerebellum vanuit verschillende sensorische centra gelegen in ruggenmerg en lage hersenstam (via ped cbl inf) en vanuit de grote hersenen via de pons (en ped cbl med).
De afvoerende (= uitgaande, = efferente) informatie van het cerebellum verloopt via de pedunculus cerebellaris superior naar het ventrolaterale deel van de thalamus (VL). Dit thalamusdeel onderhoudt reciproke verbindingen met de primaire motorschors en staat daarom ook wel bekend als motorthalamus.
Andere premotor gebieden (gebieden met ‘upper motor neurons) die informatie ontvangen vanuit de pedunculus cerebellaris superior zijn:
Nucleus ruber (rubrospinale baan)
Vestibulaire kernen (oorsprong fasciculus longitudinalis medialis)
Colliculus superior (waarom is dit ook een premotor gebied?)
Reticulaire formatie
b. In een dergelijke ‘closed loop’ zal een bepaalde groep Purkinje cellen, laten we zeggen A genaamd, informatie sturen naar een klein beperkt deel van de cerebellaire kernen (A’), die op hun beurt informatie sturen naar een bepaald deel van de thalamus (A’’), die dan weer in contact staat met een specifiek deel van de cerebrale schors (A’’’). Dat deel van de cerebrale schors heeft dan weer verbindingen met een bepaald deel van de pons (A’’’’) dat op zijn beurt dan weer verbindingen verzorgt naar dat deel van het cerebellum waar ook de groep Purkinjecellen A is gelegen. Dan is de ‘loop’ weer rond. Bij een volledig ‘open loop’ komt de informatie die vanuit het cerebellum niet meer terug bij de oorsprong. Dit soort loops kunnen natuurlijk ook een combinatie van deels ‘closed’ en deels ‘open’ zijn. Informatie komt dan deels weer terug maar is ook in staat om andere circuits te bereiken.
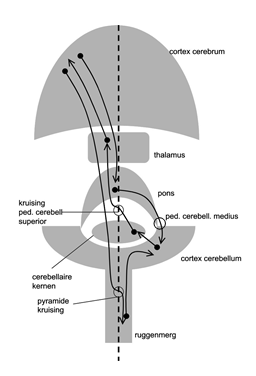
Aanvoerende verbindingen vanuit het ruggenmerg bereiken over het algemeen ongekruist het cerebellum (dus rechts blijft rechts).
De pedunculus cerebellaris superior (bevat output cerebellaire kernen) kruist in het mesencephalon (dus rechts gaat naar links).
De verbindingen tussen thalamus en motorcortex kruisen niet (dus links blijft links).
De verbindingen tussen cortex cerebri naar pons zijn ongekruist, maar die tussen pons en cerebellum zijn weer wel gekruist (dus links gaat weer naar rechts).
De corticospinale baan is wel gekruist (kruising piramidebaan) (dus links gaat weer naar rechts).
Dus rechter deel cerebellum is vooral verbonden met rechter deel lichaam en met linker cerebrum (zie verloop van pijlen in onderstaand schematische figuur). De pijlen geven hierin de richting aan die informatie (= actiepotentialen) volgt van cellichaam (zwart rondje) naar de axonuiteinden (pijlpunt).
Mosvezels eindigen in de korrelcellaag op korrelcellen. Iedere korrelcel heeft als axon een parallel vezel.
Klimvezels eindigen op Purkinje cellen. Iedere Purkinje cel ontvangt informatie va slechts één klimvezel, maar van vele tienduizenden parallelvezels.
Informatie divergeert dus het meest via het mosvezel-parallelvezelsysteem. Dit houdt in dat dezelfde informatie (van b.v. een enkele mosvezel) zeer veel Purkinjecellen bereikt. Informatie verstuurt door een enkele klimvezel bereikt slecht een enkele Purkinje cel.
Purkinje cel
Purkinje cel dendriet met spines
Korrelcellen
Klimvezel
Mosvezel rosetten
Purkinje cellen
klimvezel
pedunculus cerebellaris inferior
oliva inferior
parallelvezel
korrelcellen
moleculaire laag
korrelcellaag
pedunculus cerebellaris media
mosvezel
cerebellaire kern
pedunculus cerebellaris superior
pons
zie figuur 19.10 van Purves
collateralen klimvezelsysteem: excitatoir
collateralen mosvezelsysteem: excitatoir
Purkinje cel axonen: inhibitoir
Stercellen (stellate cells); korfcellen (basket cells); Golgi cellen.
Alle interneuronen van de cerebellaire schors zijn inhibitoir. Zie afbeelding19.10 van Purves en tekst op pp. 411 van Purves.
Het betreft een rechteroog. Dit is te zien aan de aanhechting van de m. obliquus superior die via de trochlea (katrol) die zich mediaal in de orbita bevindt, naar de oogbol loopt. De m. obliquus inferior (loopt onder het oog) heeft eveneens zijn oorsprong aan de mediale wand van de orbita.
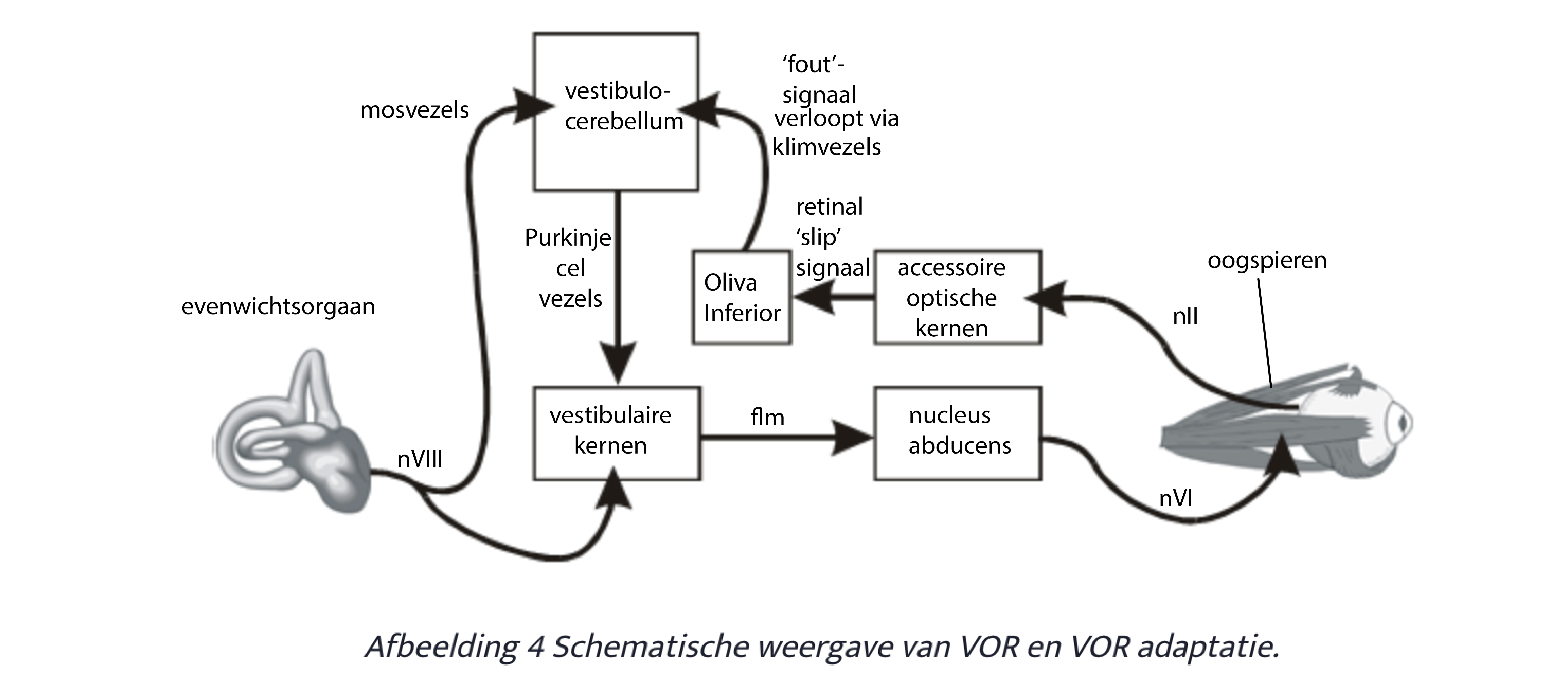
De VOR verloopt in principe via het drie-neuron-circuit: vestibulair ganglioncel - vestibulaire kerncel - motoneuron oogspier. De ‘gain’ in dit circuit (dus de mate van activatie van motoneuronen bij een bepaalde input vanuit het vestibulair orgaan, wordt mede bepaald door de input vanuit het vestibulocerebellum (via de inhibitoire input van de daar gelegen Purkinje cellen). Het vestibulocerebellum ontvangt naast directe vestibulaire input ook, via het klimvezel systeem, input over een niet- perfecte stabilisatie van het visuele veld (alleen dan wanneer de VOR niet perfect is afgesteld). Onder invloed van deze zgn. foutmeldingen zal het vestibulocerebellum zijn output
(= input van de vestibulaire kernen) veranderen totdat er geen foutmeldingen meer optreden via het klimvezel systeem. Op dat moment is de VOR perfect geijkt. Totdat, b.v. door het opzetten van een bril, of slapper worden van de oogspieren, de gain van de VOR niet meer optimaal is, er opnieuw foutsignalen worden opgewekt, en de gain in de loop van vestibulair ganglion, via vestibulocerebellum naar vestibulaire kernen zal veranderen, wat weer zal resulteren in een aangepaste VOR, etc.
Bij een geconditioneerde reflex (soms ook wel associatieve reflex genoemd) wordt een oorspronkelijk niet tot een reflex leidende stimulus (conditioning stimulus: CS) in verband gebracht met een altijd tot een reflex leidende stimulus (unconditioned stimulus: US), zodanig dat, na een voldoende aantal simultane aanbiedingen van CS en US, de CS alleen tot een reflex aanleiding geeft.
Als voorbeeld: Pavlov reflex (hond produceert speeksel bij horen bel)
Oogknipreflex, na aanbieding luchtpufje op de cornea als US en toon als CS. Na verloop van herhaaldelijk aanbieden van US samen met CS zal een reflex optreden na aanbieding van een CS zonder US.
I. Het cerebellum kan blijkbaar een veelheid van mogelijke CS signalen koppelen aan een US. Dit is alleen mogelijk als al die CS signalen grote delen van de cerebellaire schors bereikt. Dit is mogelijk via het sterk divergerende karakter van het mosvezel-parallel vezel systeem.
Motorische problemen uitten zich veel eerder in het dagelijks leven dan veelal vage autonome en/of cognitieve klachten.
Cerebellaire ataxie wordt waarschijnlijk veroorzaakt doordat de cerebellaire controle over nauwkeurig aangestuurde bewegingen en reflexen niet meer functioneert. De balans tussen allerlei ‘geleerde’ en ‘geautomatiseerde’ bewegingen valt weg (vergelijk met VOR adaptatie). Bewegingen vallen daarom veelal te groot of te klein uit.
De cerebellaire schors is veel groter dan de kernen. De gehele schors convergeert dus op het cerebellaire kern gebied. Dit houdt in dat meerdere schors delen op hetzelfde kerngebied zullen projecteren. Het is niet waarschijnlijk dat die allemaal tegelijk aangedaan zullen zijn. De kernen functioneren daarentegen als output station.
Laesie van een (deel van) de kernen, betekent dus disfunctioneren van een groot deel van de cerebellaire schors.
Wanneer belangrijke aanvoerende systemen voor het cerebellum aangedaan zijn zoals de corticopontine baan of spinocerebellaire verbindingen kan het cerebellum zijn taken niet goed uitvoeren en zullen er cerebellaire disfuncties kunnen ontstaan die zich vooral zullen resulteren in ataxie (= ‘problemen’ met coördineren van bewegingen, zie tekst Purves blz 4–414-416).